- Специфическая соматическая рекомбинация
-
Специфическая соматическая рекомбинация (V(D)J рекомбинация, англ. V(D)J recombination) — особый процесс сайт-специфической генетической рекомбинации, происходящий в иммунных клетках у позвоночных животных, который случайным образом переставляет и перестраивает гены, ассоциированные со специфическими белками, играющих важную роль в иммунном ответе[1]. В результате этого процесса генерируется разнообразный набор молекул Т-клеточных рецепторов (TCR, ТкР) и иммуноглобулинов (Ig), ответственных за распознавание антигенов как чужеродного происхождения бактерий, вирусов и других инфекционных агентов, так и эндогенных антигенов, таких как например клетки злокачественных опухолей.[2]
Содержание
Введение
Иммуноглобулины человека (точно также как и В-клеточные рецепторы) состоят из лёгких и тяжёлых цепей, и те и другие содержат консервативные (С) и вариабельные (V) участки и кодируются нижеследующими тремя локусами:
- Локус тяжёлой цепи (IGH@), локализированный на хромосоме 14, кодирующий тяжёлые цепи иммуноглобулинов
- Каппа (κ)-локус (IGK@) , расположенный на хромосоме 2, кодирующий лёгкие цепи иммуноглобулинов
- Лямбда (λ)-локус (IGL@) , расположенный на хромосоме 22, кодирующий лёгкие цепи иммуноглобулинов
Гены вариабельных регионов многочисленны, их можно разделить на три типа последовательностей. Так, регион ассоциированный с тяжёлой цепью иммуноглобулина содержит 123 VH сегментов, из которых 79 являются псевдогенами и 44 содержат открытые рамки считывания, 27 DH и 6 JH генов[3]. Регионы, ответственные за лёгкие цепи также содержат V и J последовательности, но не содержат D-последовательностей. Таким образом, возможно большое количество комбинаций, что порожает великое разнообразие иммуноглобулинов.
Большинство Т-клеточных рецепторов содержат альфа- и бета-цепи. Гены Т-клеточных рецепторов сходны с генами иммуноглобулинов в том, что они также содержат множественные V, D и J гены в β-цепи (и V и J гены в α-цепи), которые перегруппируются в процессе дифференциации тимоцита с целью обеспечения уникального рецептора антигена.
Иммуноглобулины
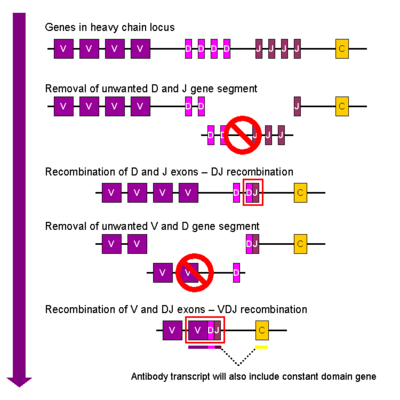 Упрощённая схема of V(D)J рекомбинации
Упрощённая схема of V(D)J рекомбинации
Тяжёлые цепи
В процессе созревания В-клетки происходит рекомбинация между одним D сегментом и одним J сегментом. ДНК между этими двумя последовательностями удаляется. За D-J рекомбинацией следует присоединение одного V сегмента из области, близкой к 5' концу от новообразованного DJ комплекса, в результате образуется перегруппированный VDJ ген. Все последовательности между V и D сегментами удаляются из генома клетки. С последовательности новообразованного VDJ гена синтезируется пре-мРНК, как μ-цепи, так и δ-цепи (Cμ и Cδ) (т.е. первичный транскрипт имеет структуру V-D-J-Cμ-Cδ). Затем происходит процессинг пре-мРНК, заключающийся в полиаденилировании 3'-конца транскрипта и вырезании последовательности между VDJ и С сегментом. Трансляция новообразовавшейся мРНК приводит к синтезу тяжёлой цепи Ig μ.
Лёгкие цепи
κ- и λ-цепи иммуноглобулинов формируются весьма сходным образом за исключением отсутствия D сегмента. Таким образом, первая стадия рекомбинации заключается в объединении V и J сегментов с образованием VJ комплекса до добавления последовательности консервативного региона во время транскрипции. Трансляция процессированной РНК приводит к синтезу Ig κ либо Ig λ лёгких цепей.
Сборка тяжёлых и лёгких цепей приводит к мембранносвязанной форме иммуноглобулина IgM, который экспрессируется на поверхность зрелой В-клетки.
Т-клеточные рецепторы
В процессе созревания тимоцитов цепи гены Т-клеточного рецептора проходят сходную последовательность рекомбинаций и перестроек что и гены иммуноглобулинов. Первой происходит D-J рекомбинация в β-цепи ТкР. При этом происходит либо объединение одного Dβ1 сегмента с одним из шести Jβ1 сегментов либо объединение Dβ2 сегмента с одним из семи Jβ2 сегментов. Вслед за D-J рекомбинацией спроисходит Vβ-DβJβ перегруппировка. Все последовательности между Vβ-Dβ-Jβ генами удаляются и новообразованная последовательность транскрибируются с присоединением консервативного участка с образованием Vβ-Dβ-Jβ-Cβ мРНК транскрипта с вырезанием всех междулежащих последовательностей с трансляцией в полноценную TCR Cβ цепь
Перестройка α-цепи аналогична таковой в лёгких цепях иммуноглобулинов. Объединение α- и β- цепей приводит к формированию полноценного αβ-TCR рецептора, экспрессируемого большинством Т-клеток.
Механизм
Сигнальные последовательности
V, D и J последовательности фланкируются специальными сигнальными последовательностями - англ. recombination Signal Sequences (RSS), распознающихся специальными ферментами, т.н. VDJ рекомбиназами. RSS содержат консервативную последовательность семи нуклеотидов (гептамер), находящуюся после кодирующей последовательности и следующим за ней спейсером, содержащим неконсервативную последовательность длиной либо 12, либо 23 нуклеотида, и следующим консервативным нонамером (англ. nonamer, т.е. 9-членная последовательность), содержащим 9 нуклеотидов. RSS расположены на 3' V сегмента и на 5' J сегмента, т.е. на участках, вовлечённых в присоедниении последовательностей друг к другу. Рекомбинируют только пары разных спейсерных RSS (т.е. 12-членный спейсер будет рекомбинировать только с 23-членным). Это т.н. правило рекомбинации 12/23
VDJ рекомбиназа
Под VDJ рекомбиназой понимают комплекс ферментов, как лимфоцит-специфичных, так и не специфичных для какого-либо определённого типа клеток. Первая стадия процесса происходит с участием важных лимфоцит-специфических ферментов - RAG1 и RAG2 (англ. Recombination activating gene, en:Recombination activating gene). Эти гены образуют комплекс, узнающий RSS и индуцирующий вырезание ДНК в RSS-последовательностях. Вырезание происходит только в одной цепи ДНК, что приводит к образованию шпильки.
Другие ферменты VDJ рекомбиназного комплекса не являются лимфоцит-специфическими и производят репарацию ДНК вслед за действием RAG1 и RAG2 ферментов. Один из этих ферментов это ДНК-зависимая протеинкиназа, которая производит репарарацию двуцепочечной ДНК. ДНК-зависимая протеинкиназа присоединяется к обоим концам повреждённой ДНК и образует комплекс с другими белками, в т.ч. и с нуклеазой Artemis — XRCC4 (англ. X-ray repair cross-complementing factor 4), ДНК лигазой IV, белком Cernunnos (также называемым XLF или XRCC4-подобным фактором) и несколькими ДНК-полимеразами. ДНК-зависимая протеинкиназа фосфолирирует каждый из концов ДНК, тем самым активируя нуклеазу Artemis. Нуклеаза Artemis затем разрезает шпильку, образованную RAG ферментами[4]. Белки XRCC4 и Cernunnos работают в взаимосвязи с ДНК-зависимой протеинкиназой, выравнивают образовавшиеся концы ДНК друг с другом, а также помогают работе фермента терминальной дезоксинуклеотидилтрансферазы, которая случайным образом присоединяет нуклеотиды к открытым концам ДНК и образуя т.н. липкие концы, приводя к т.н. стыковому разнообразию (англ. Junctional diversity, en:Junctional diversity). ДНК полимеразы λ и μ присоединяют нуклеотиды, необходимые для комплиментарности липких концов друг другу. Лиагаза IV окончательно сшивает липкие концы друг с другом, завершая процесс присоединения сегментов друг к другу [5].
Из-за большого разнообразия сайтов для нуклеазы Artemis, а также случайного добавления нуклеотидов терминальной нуклеотидилтрансферазой, окончательные последовательности ДНК, ассоциированные с иммуноглобулинами являются чрезвычайно вариабельными, даже когда одни и те же V, D, или J присоединяются друг к другу. В результате VDJ рекомбинации формируются иммуноглобулины к антигенам, с которыми никогда не сталкивались ни сам организм, ни его предки.
Примечания
- ↑ Janeway CA, Jr. et al.' Immunobiology.. — 6th. — Garland Science, 2005. — ISBN 0-443-07310-4
- ↑ Abbas AK and Lichtman AH Cellular and Molecular Immunology. — 5th. — Saunders, Philadelphia, 2003. — ISBN 0-7216-0008-5
- ↑ Li A, Rue M, Zhou J, et al. (June 2004). «Utilization of Ig heavy chain variable, diversity, and joining gene segments in children with B-lineage acute lymphoblastic leukemia: implications for the mechanisms of VDJ recombination and for pathogenesis». Blood 103 (12): 4602–9. DOI:10.1182/blood-2003-11-3857. PMID 15010366.
- ↑ Y. Ma, U. Pannicke, K. Schwarz and M.R. Lieber (2004). «Hairpin opening and overhang processing by an Artemis/DNA-dependent protein kinase complex in nonhomologous end joining and V(D)J recombination». Cell 108 (6): 781–794. DOI:10.1016/S0092-8674(02)00671-2. PMID 11955432.
- ↑ D.C. van Gent and M. van der Burg (2007). «Non-homologous end-joining, a sticky affair». Oncogene 26 (56): 7731–7740. DOI:10.1038/sj.onc.1210871. PMID 18066085.
Литература
- Hartwell LH, Hood L, Goldberg ML, Reynolds AE, Silver LM, Veres RC Chapter 24, Evolution at the molecular level. In: Genetics. — New York: McGraw-Hill, 2000. — P. 805–807. — ISBN 0072995874
- V(D)J Recombination. Series: Advances in Experimental Medicine and Biology, Vol. 650 Ferrier, Pierre (Ed.) Landes Bioscience 2009, XII, 199 p. ISBN 978-1-4419-0295-5
Иммунная система / Иммунология Системы Адаптивная иммунная система и Врожденная иммунная система · Гуморальная иммунная система и Клеточная иммунная система · Система комплемента (Анафилотоксины) · Intrinsic immunity Антигены и антитела Антиген (Суперантиген, Аллерген) · Гаптены · Fab · Fc ·
Эпитоп (Линейный эпитоп, Конформационный эпитоп)
Антитела (Моноклональные антитела, Поликлональные антитела, Аутоантитела) · Polyclonal B cell response · Аллотипы антител · Изотипы антител · Идиотипы антител
Иммунный комплексКлетки иммунной системы
ЛейкоцитыЛимфоидные: Т-лимфоциты · B-лимфоциты · Естественные киллеры · Плазматические клетки
Миелоидные: Тучные клетки · Базофилы · Эозинофилы · Макрофаги
Фагоциты: Нейтрофилы · Макрофаги, Ретикулоэндотелиальная система
Антигенпредставляющие клетки: Дендритные клетки · Макрофаги · B-лимфоциты · Презентация антигенаИммунитет и толерантность действие: Иммунитет · Autoimmunity · Аллергия · Воспаление · Кросс-реактивность
бездействие: Иммунологическая толерантность (Central, Peripheral, Clonal anergy, Clonal deletion) · ИммунодефицитРецепторы Т-клеточный рецептор · Fc рецептор Иммуногенетика Соматический гипермутагенез · V(D)J рекомбинация · Класс-переключение · Главный комплекс гистосовместимости/HLA · Гистосовместимость Вещества Цитокины · Опсонин · Cytolysin Другое Диагностическая иммунология Органы иммунной системы Тимус · Селезёнка · Лимфатические узлы · Кровь · Костный мозг · Лимфа · Заболевания иммунной системы (Иммунодефицит) Категории:- Лимфоциты
- Молекулярно-генетические процессы
Wikimedia Foundation. 2010.